CAM is an adaptation for increased efficiency in the use of water, and so is typically found in plants growing in dry conditions
CAM was first suspected by Nicolas-Théodore de Saussure in 1804 in his Recherches Chimiques sur la Vegetation, confirmed and refined by Aubert, E. in 1892 in his Recherches physiologiques sur les plantes grasses.

Expounded upon by Richards, H. M. 1915, in Acidity and Gas Interchange in Cacti, Carnegie Institution. Ranson and Thomas coined the term CAM in 1940. But, they were not the first to discover this cycle.
It was observed by the botanists Ranson and Thomas in the succulent family Crassulaceae (which includes jade plants and Sedum). Its name refers to acid metabolism in Crassulaceae, not the metabolism of “crassulacean acid”.
In a plant using full CAM, the stomata in the leaves remain shut during the day. This reduces evapotranspiration. They open at night to collect carbon dioxide (CO2). The CO2 is stored as the four-carbon acid malate in vacuoles at night. Then, during the daytime, the malate is transported to chloroplasts. There, it is converted back to CO2. This CO2 is then used during photosynthesis.
The pre-collected CO2 is concentrated around the enzyme RuBisCO, increasing photosynthetic efficiency. The mechanism was first discovered in plants of the family Crassulaceae.
During the night, a plant employing CAM has its stomata open. This allows CO2 to enter. It is fixed as organic acids that are stored in vacuoles. During the day, the stomata are closed. This prevents water loss. The carbon is released to the Calvin cycle so that photosynthesis may take place.
The carbon dioxide is fixed in the cytoplasm of mesophyll cells. This is done by a PEP reaction akin to that of the C4 pathway. But, unlike the C4 mechanism, the resulting organic acids are stored in vacuoles for later use. They are not instantly passed on to the Calvin cycle.
The latter cannot function during the night. The light reactions that provide it with ATP and NADPH cannot take place.
During the day, the CO2-storing organic acids are released from the vacuoles of the mesophyll cells. These acids then enter the stroma of the chloroplasts. In the chloroplasts, an enzyme releases the CO2. This released CO2 then enters the Calvin cycle.
The most important advantage of CAM to the plant is its ability to keep most leaf stomata closed. This happens during the day.
Plants employing CAM are most common in arid environments, where water comes at a premium. These plants can keep stomata closed during the hottest and driest part of the day. This ability reduces the loss of water through evapotranspiration. It allows such plants to grow in environments that would otherwise be far too dry.
Plants using only C3 carbon fixation, for example, lose 97% of the water they uptake through the roots to transpiration. This is a high cost avoided by plants able to employ CAM.
The C4 pathway bears resemblance to CAM and acts to concentrate CO2 around RuBisCO, thereby increasing its efficiency. CAM concentrates it temporally, providing CO2 during the day, and not at night.
When respiration is the dominant reaction. C4 plants, in contrast, concentrate CO2 spatially. A RuBisCO reaction center in a “bundle sheath cell” is inundated with CO2. Due to the inactivity required by the CAM mechanism, C4 carbon fixation has greater efficiency in terms of PGA synthesis.
Plants with CAM must control storage of CO2 and its reduction to branched carbohydrates in space and time.
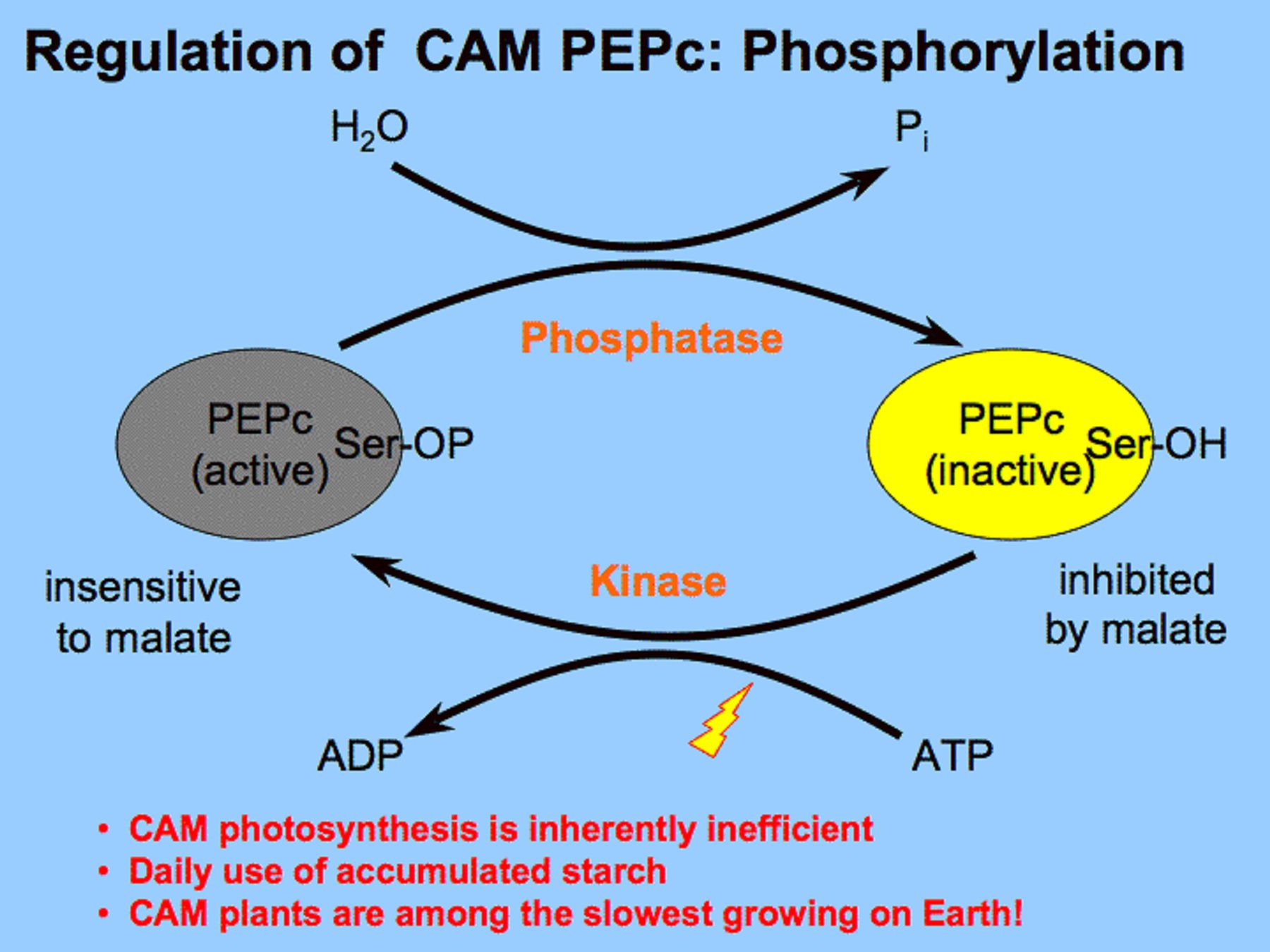
At low temperatures (frequently at night), plants using CAM open their stomata, CO2 molecules diffuse into the spongy mesophyll’s intracellular spaces and then into the cytoplasm. Here, they can meet phosphoenolpyruvate (PEP), which is a phosphorylated triose.
During this time, the plants are synthesizing a protein called PEP carboxylase kinase (PEP-C kinase). Its expression can be inhibited by high temperatures, often at daylight. The presence of malate also inhibits its expression. PEP-C kinase phosphorylates its target enzyme PEP carboxylase (PEP-C).
Phosphorylation dramatically enhances the enzyme’s capability to catalyze the formation of oxaloacetate. This oxaloacetate can then be transformed into malate by NAD+ malate dehydrogenase.
Malate is then transported via malate shuttles into the vacuole, where it is converted into the storage form malic acid. In contrast to PEP-C kinase,
PEP-C is synthesized continuously. It is almost inhibited in daylight. This happens either by dephosphorylation via PEP-C phosphatase or directly by binding malate. The latter is not possible at low temperatures since malate is efficiently transported into the vacuole. Whereas PEP-C kinase readily inverts dephosphorylation.
In daylight, plants using CAM close their guard cells and discharge malate that is subsequently transported into chloroplasts.
There, depending on plant species, it is cleaved into pyruvate and CO2 either by malic enzyme or by PEP carboxykinase. CO2 enters the Calvin cycle. It is a coupled and self-recovering enzyme system. This system is used to build branched carbohydrates.
The by-product pyruvate can be further degraded in the mitochondrial citric acid cycle. This degradation provides extra CO2 molecules for the Calvin Cycle. Pyruvate can also be used to recover PEP through pyruvate phosphate dikinase. This is a high-energy step. It requires ATP and an additional phosphate.
During the following cool night, PEP is finally exported into the cytoplasm. There, it is involved in fixing carbon dioxide via malate.
Plants use CAM to different degrees. Some are “obligate CAM plants”, i.e. they use only CAM in photosynthesis. However, they vary in the amount of CO2 they are able to store as organic acids. They are sometimes divided into “strong CAM” and “weak CAM” plants on this basis.
Other plants show “inducible CAM”. They can switch between using either the C3 or C4 mechanism and CAM depending on environmental conditions.
Another group of plants employs “CAM-cycling”. In this process, their stomata do not open at night. Instead, the plants recycle CO2 produced by respiration. They also store some CO2 during the day.
Plants showing inducible CAM and CAM-cycling are typically found in conditions where periods of water shortage alternate. They also occur when periods when water is freely available. Periodic drought is one cause of water shortage. Plants that grow on trees or rocks (as epiphytes or lithophytes) also experience variations in water availability.
Salinity, high light levels, and nutrient availability are other factors that have been shown to induce CAM.
Since CAM is an adaptation to arid conditions, plants using CAM often display other xerophytic characters. These include thick, reduced leaves with a low surface-area-to-volume ratio and a thick cuticle. They also have stomata sunken into pits. Some shed their leaves during the dry season; others (the succulents) store water in vacuoles.
CAM also causes taste differences. Plants have an increasingly sour taste during the night. They become sweeter-tasting during the day. Malic acid is stored in the vacuoles of the plants’ cells at night. It is used up during the day.
The majority of plants possessing CAM are either epiphytes (e.g., orchids, bromeliads) or succulent xerophytes (e.g., cacti, cactoid Euphorbias).
Additionally, CAM is found in hemiepiphytes (e.g., Clusia), lithophytes (e.g., Sedum, Sempervivum), terrestrial bromeliads, and wetland plants.
There is also one halophyte, Mesembryanthemum crystallinum, one non-succulent terrestrial plant, (Dodonaea viscosa) and one mangrove associate (Sesuvium portulacastrum).
Some plants can switch between different methods of carbon fixation. For example, Portulacaria afra is better known as Dwarf Jade Plant. This plant normally uses C3 fixation but can use CAM if it is drought-stressed. Another plant is Portulaca oleracea, better known as Purslane. It normally uses C4 fixation but is also able to switch to CAM when drought-stressed.
CAM has evolved convergently many times. It occurs in 16,000 species. This signifies about 7% of plants. These species belong to over 300 genera and around 40 families. Still, this is thought to be a considerable underestimate.
It is found in quillworts (relatives of club mosses). It is also found in ferns and in Gnetopsida. Yet, the great majority of plants using CAM are angiosperms (flowering plants).
CAM photosynthesis is also found in aquatic species in at least four genera. These include Isoetes, Crassula, Littorella, Sagittaria, and possibly Vallisneria. This adaptation is seen in various species, such as Isoetes howellii and Crassula aquatica.
These plants follow the same nocturnal acid accumulation and daytime deacidification as terrestrial CAM species.
The reason for CAM in aquatic plants is not due to a lack of available water. It is due to a limited supply of CO2. CO2 is limited due to slow diffusion in water, 10000x slower than in air.
The problem is especially acute under acid pH. In this condition, the only inorganic carbon species present is CO2. There is no available bicarbonate or carbonate supply.
Aquatic CAM plants capture carbon at night. Carbon is abundant at night due to a lack of competition from other photosynthetic organisms. This also results in lowered photorespiration due to less photosynthetically generated oxygen.
Aquatic CAM is most marked in the summer months. There is increased competition for CO2 during this period compared to the winter months. Yet, in the winter months, CAM still has a significant role.
Like what you read? Appreciate the information. We are a non-profit 501c3! Thank us with a contribution by clicking DONATE!
Leave a Reply